Emai:marketing@yakkaa.com
業務谘詢專線:400-780-8018
Tel: +1(626)986-9880(U.S. - West Coast)
0044 7790 816 954 (Europe)
Email: marketing@medicilon.com
地址:上海市浦東新區川大路585號
郵編:201299
電話:+86 (21) 5859-1500(總機)
傳真:+86 (21) 5859-6369





© 2023 上海hjc黄金城生物醫藥股份有限公司 保留所有權利 滬ICP備10216606號-3
滬公網安備 31011502018888號 | 網站地圖











業務谘詢
中國:
Email: marketing@yakkaa.com
業務谘詢專線:400-780-8018
(僅限服務谘詢,其他事宜請撥打川沙總部電話)
川沙總部電話: +86 (21) 5859-1500
海外:
+1(626)986-9880(U.S. - West Coast)
0044 7790 816 954 (Europe)
Email:marketing@medicilon.com




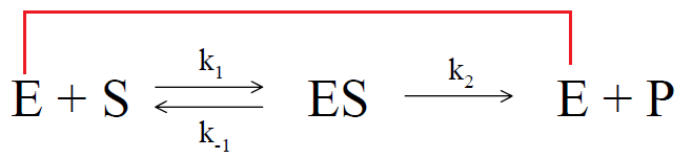
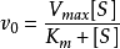
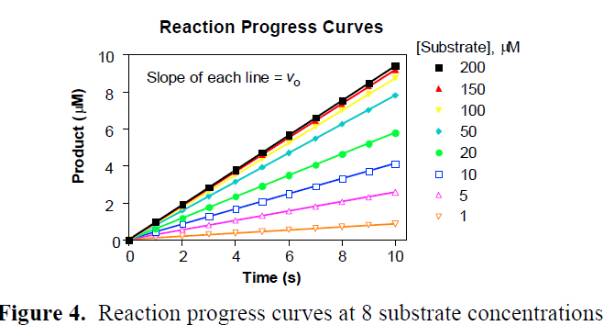
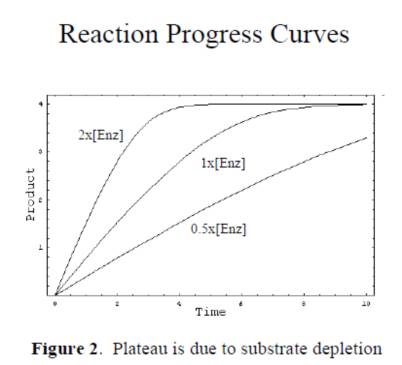
1.必須是動力學讀數,選取初始反映的線性段的斜率作為初始反應的速度。終點法讀數的結果很多時候並不可靠,應為酶催化反應隨著時間的進行底物的消耗,反應速度會從勻速到非均速變化,如果采用終點法可能得到不真實的初始速度,如下圖:

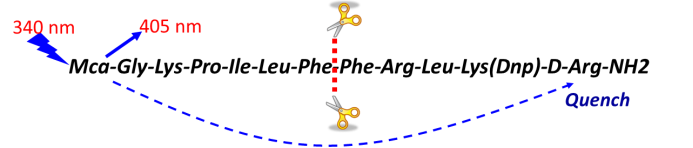
底物的選擇和配套的檢測方式,如下圖某蛋白水解酶,其識別的位點是氨基酸線性表位,可以人工合成多肽,在其N端和C端分別加入熒光基團和淬滅基團,正常狀態下或者酶活抑製狀態,該多肽在酶標儀340nm激發時,其發射光會被淬滅基團吸收,在405nm無信號產生;酶催化反應會將該多肽剪切,當酶標儀340nm激發時,405nm有信號產生。
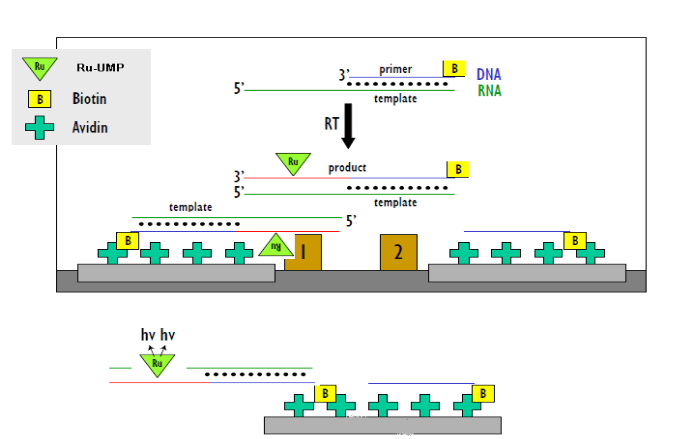
又如某病毒逆轉錄酶,合成RNA模板和對應引物,在其引物上進行生物素標記,其中堿基上進行Ru標記,隨著酶催化的逆轉錄反應的進行,引物上不斷有Ru標記堿基上去形成核苷酸序列,通過親和素的板子捕捉引物,檢測Ru信號值來評判酶的催化和抑製的活性。
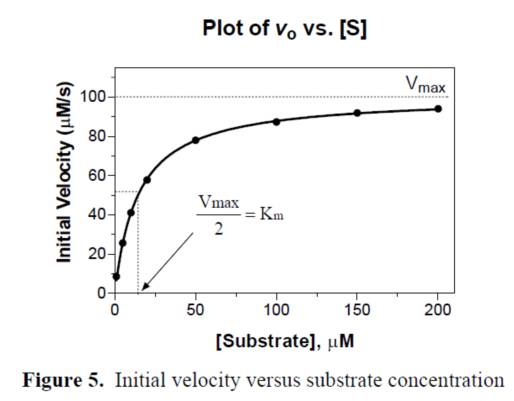
當酶對應的底物確定以後,選取少量的酶濃度(pM或者nM)對底物進行Km測定,選取底物的Km濃度作為篩選的反應濃度,選取該濃度下線性的反應時間內的某個時間點作為反應終止或者檢測的時間。酶和梯度稀釋的化合物進行預孵育後,加入Km濃度的底物,反應一段時間後進行檢測,這個就是一般酶學篩選模型。
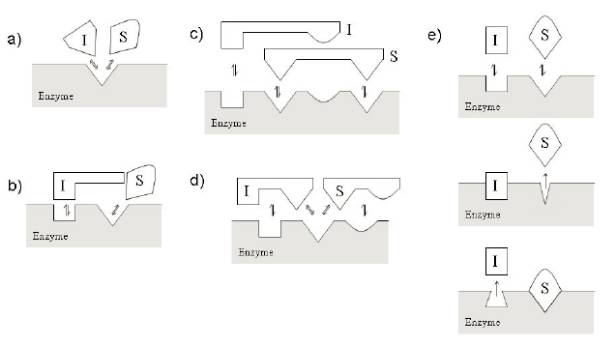
competitive競爭性抑製劑隻和自由的酶結合,通常情況下和底物共同競爭結合位點(活性中心),當然也有競爭性抑製劑和底物互斥的情況出現,兩個中隻有一個能夠和自由的酶結合。在競爭性抑製劑存在的情況下,酶相對於底物的Km和Vmax測定時,測得得表觀Km值增加,Vmax值不變,如下圖所示,幾種競爭性抑製劑的圖示:
非競爭性抑製劑可以和自由的酶結合,也可以和酶-底物複合物結合,抑製劑結合的位點和活性中心位點不在相同的表位,最終也會導致酶催化活性的喪失。在非競爭性抑製劑存在的情況下,酶相對於底物的Km和Vmax測定時,測得得表觀Km值不變,Vmax值降低,如下圖所示:

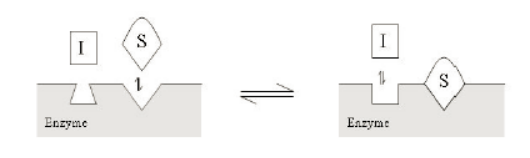
不競爭性抑製劑隻和酶-底物複合物結合,形成失活的抑製劑-酶-底物複合物。酶相對於底物的Km和Vmax測定時,測得得表觀Km值降低,Vmax值降低。如下圖所示:
Ki描述的是抑製劑和酶的結合強弱,是抑製劑和酶的解離平衡常數,是個狀態函數,不會受到反應體係中底物濃度的影響,不同實驗得到的Ki值具有可比性;而IC50則是在特定酶學檢測方法中酶活抑製一半時抑製劑的濃度,根據不同類型的抑製劑,底物濃度可能對IC50產生影響,不同實驗得到的IC50值不具可比性。根據不同類型的抑製劑,在理想條件下(酶濃度遠遠小於底物濃度進行反應),Ki和IC50的關係如下:

不同抑製劑類型IC50和反應體係中加入的底物濃度的關係示意圖如下:

 相關新聞
相關新聞川沙總部
地址: 上海市浦東新區川大路585號
郵編: 201299
電話: +86 (21) 5859-1500(總機)
傳真: +86 (21) 5859-6369
海外:
Email: marketing@medicilon.com
Tel: +1(626)986-9880(U.S. - West Coast)
Tel: 0044 7790 816 954 (Europe)
Tel: +82 70-8269-5849 (Korea)
Tel: +81 80-4421-6898 (Japan)


 關於我們
關於我們